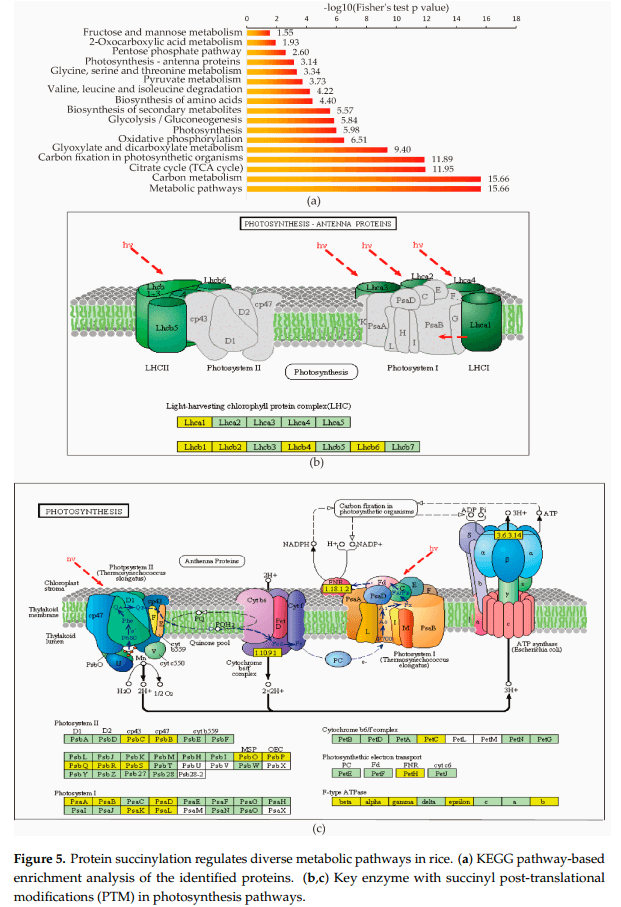
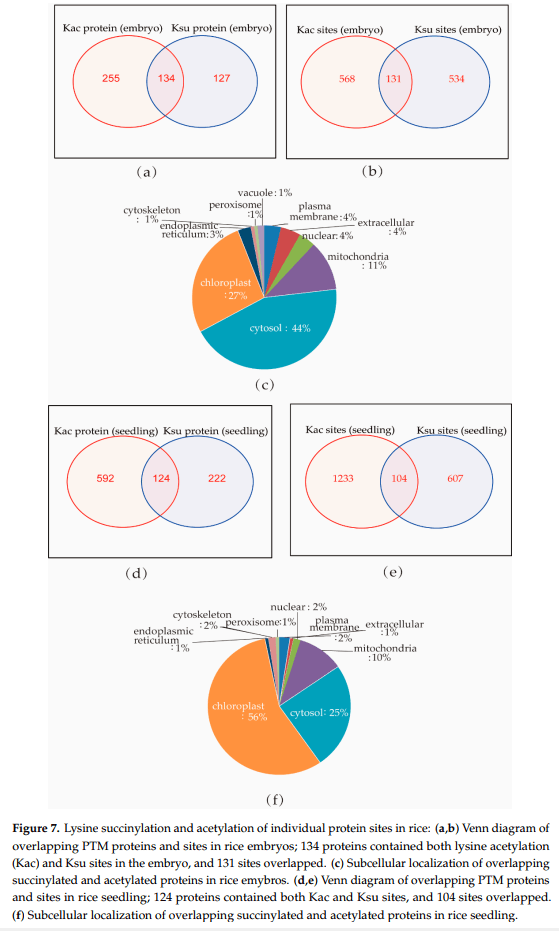
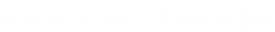作者发现在许多与光合作用相关的蛋白质亚基中存在琥珀酰化,如采光叶绿素蛋白复合物(LHC)中的Lhca1、Lhcb1、Lhcb2、Lhcb4和Lhcb6(图5B),以及光系统I I中的PsbC、PsbB、PsbO、PsbP、PsbQ、PsbR、PsbS、PsaA、PsaB、PsaD、PsaK、PsaL,细胞色素b6/f复合物中的PetC,光系统电子传递和β、α、γ、epsilonf型atp酶中的PetH(图5C)。赖氨酸琥珀酰化也参与碳代谢,包括碳固定。同时发现赖氨酸琥珀酰化几乎发生在Calvin循环的每个分支中:许多蛋白质,如果糖-1,6-双磷酸酶、磷酸甘油酸激酶和磷酸布洛激酶,被鉴定为琥珀酰化蛋白质。