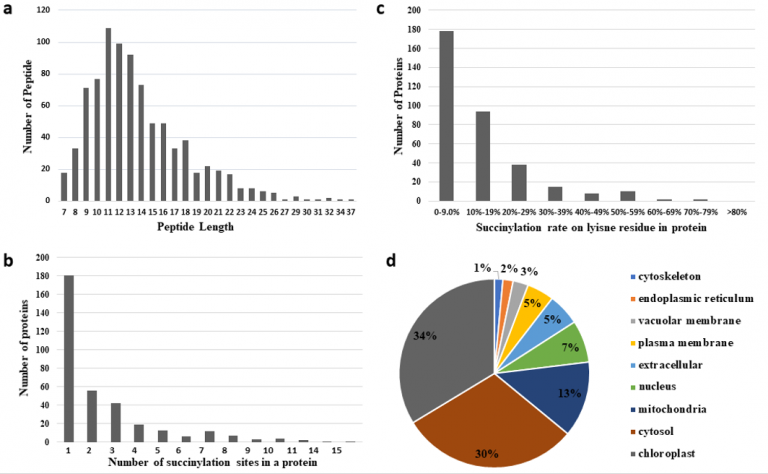
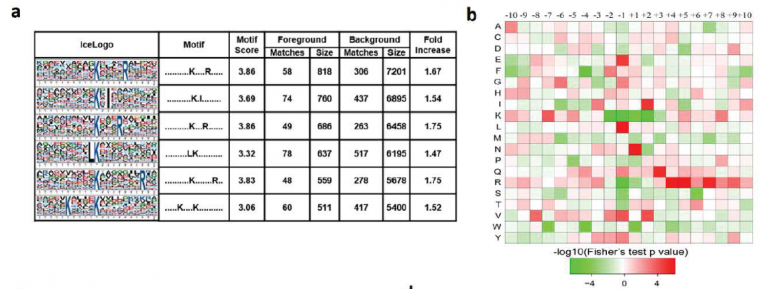
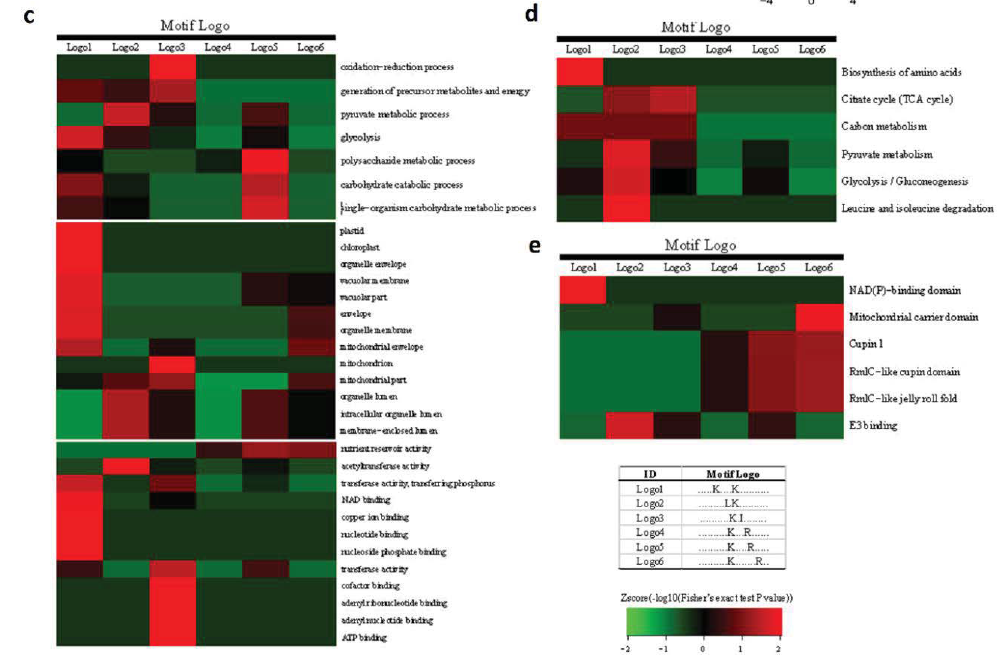
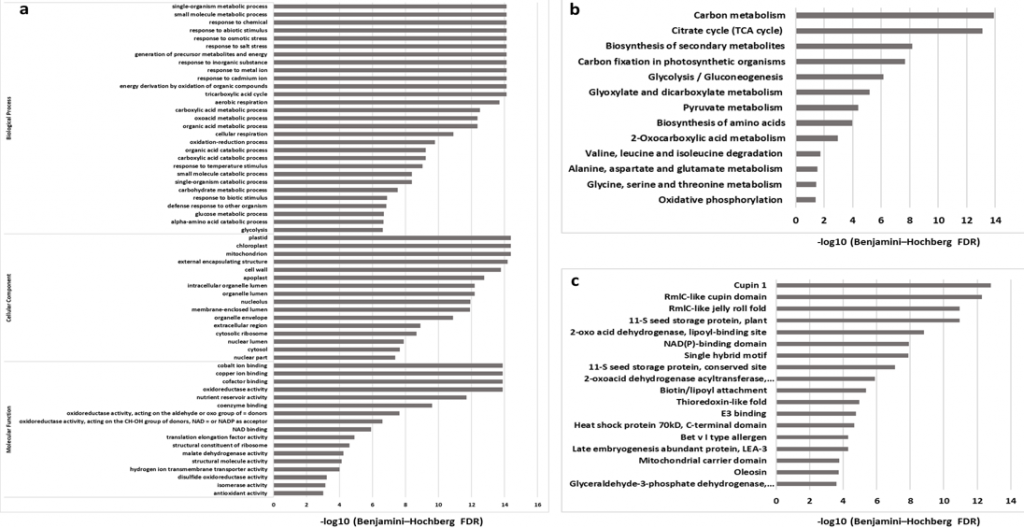
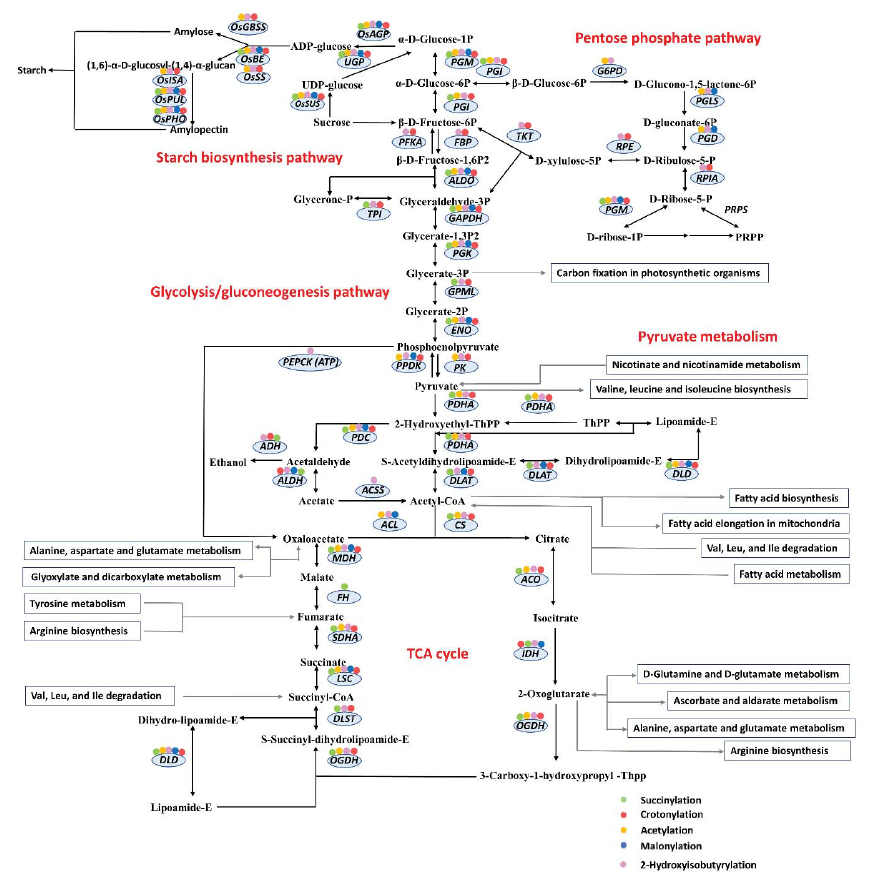
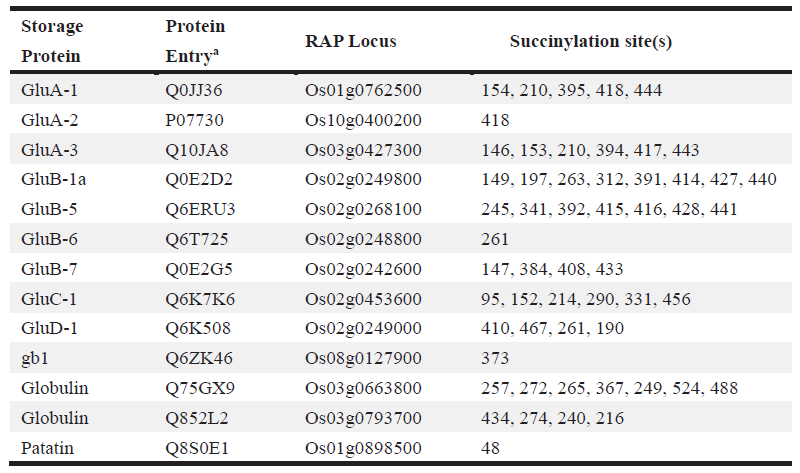
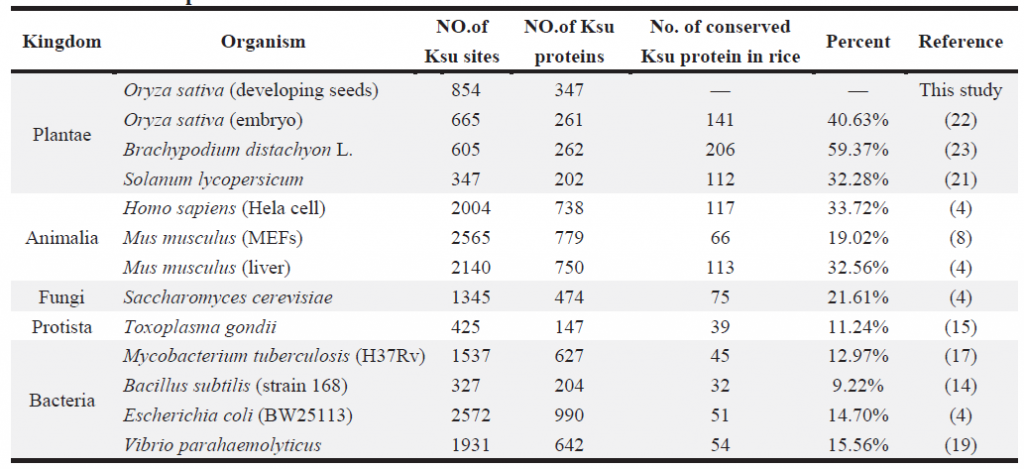
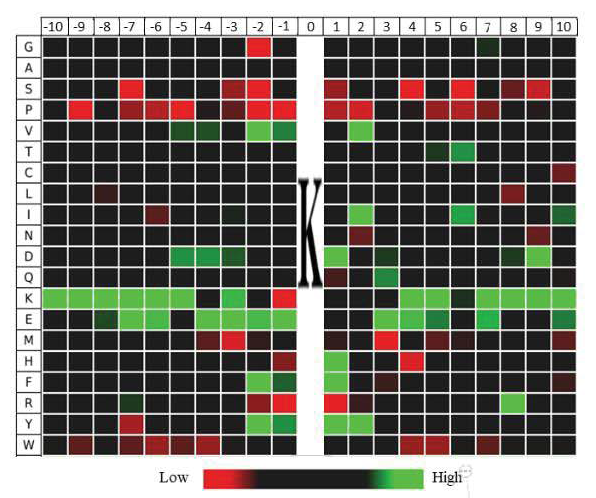
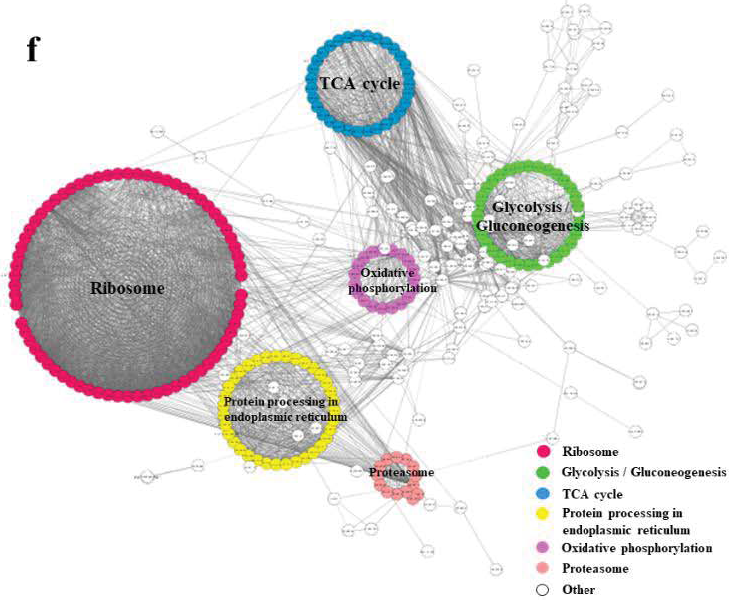
2. 水稻发育种子总体乙酰化修饰水平评估
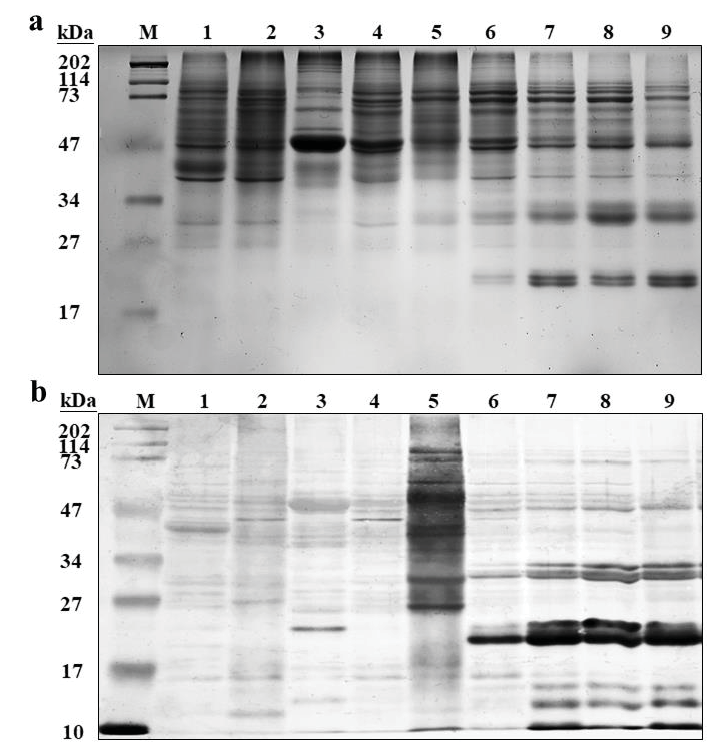
琥珀酰化修饰依赖succinyl-CoA和pH。对开花后灌浆中的水稻种子进行泛琥珀酰化抗体WB,发现在对应pH下体外succinyl-CoA高含量会提升蛋白的琥珀酰化修饰水平(lane1 样品蛋白、lane2 蛋白+PBS、lane3 蛋白+PBS+0.5mM succinyl-CoA、lane4 蛋白+PBS+1mMsuccinyl-CoA,上图SDSPAGE,下图WB结果)。
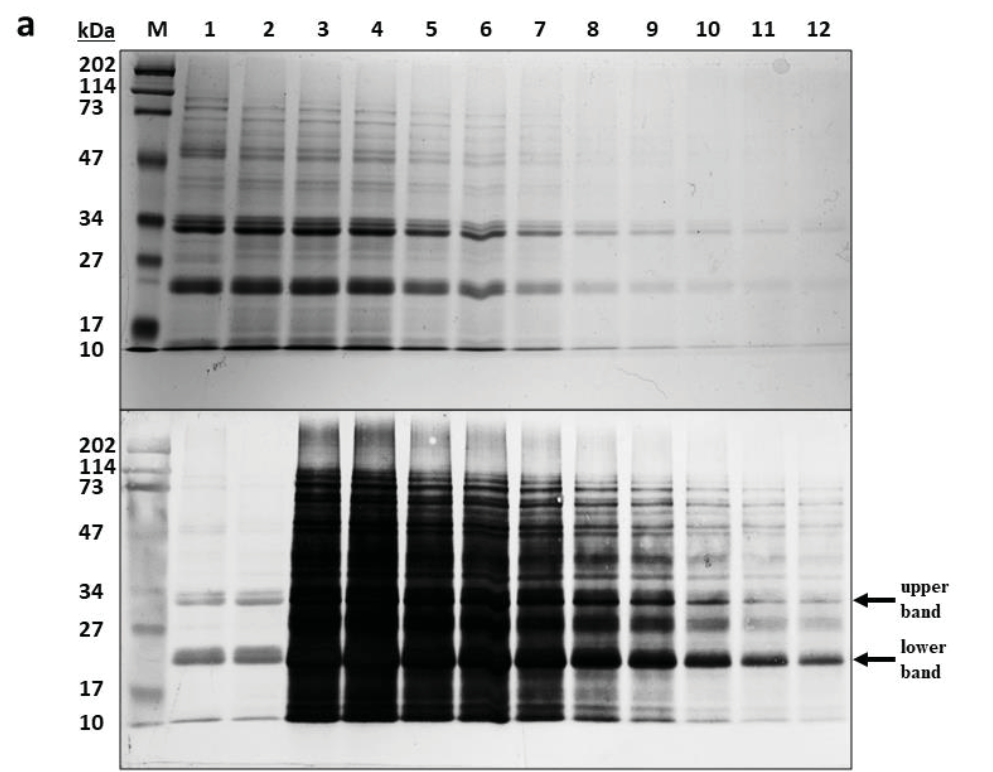
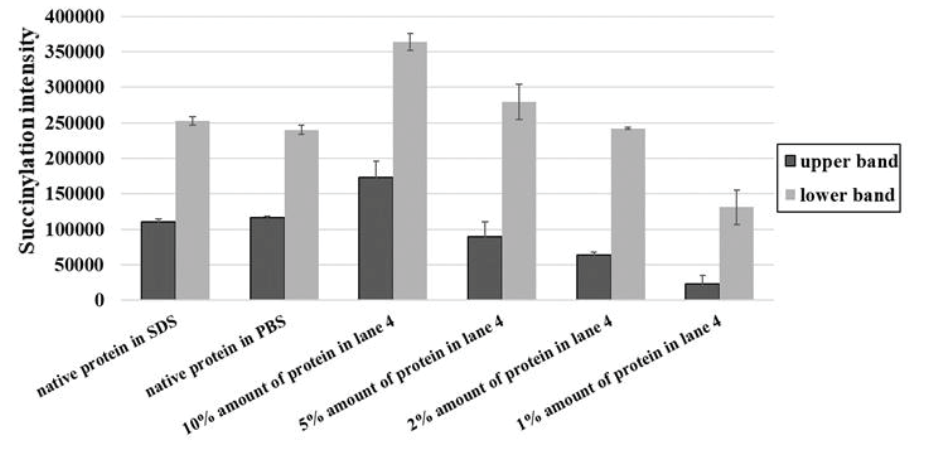
在保持相同PBS和succinyl-CoA浓度下,依次降低样品蛋白上样量(lane 4-12),发现两条主带的WB响应浓度随着蛋白总量的下降而持续下降。通过WB定量分析,灌浆期水稻种子的琥珀酰化修饰占有率在2%-10%之间。