
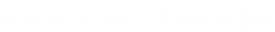
文献解读
项目论文|不同土豆品种对致病疫霉抗性的TMT标记定量比较蛋白质组学研究-International Journal of Molecular Sciences-201901
为了更好地了解马铃薯(cv.Sarpo Mira)过程中蛋白质的相互作用–疫霉互作不亲和,作者研究了cv.Sarpo Mira的蛋白质组动力学。在接种后0小时(对照)、48小时(EI)和120小时(LI)三个关键时间点,用标记定量蛋白质组学(TMT)将虫害分离物的游动孢子悬浮液叶面施用后,分为早期和晚期。共鉴定出1229个差异表达蛋白(DEPs)。以Sarpo-Mira配对比较两个疾病分期,包括共同共病期、特定病期分别在早期和晚期。80%以上的蛋白质丰度变化在感染早期上调,而更多的DEPs(61%)在感染后期下调。表达模式、功能分类和富集试验强调了细胞壁相关防御反应蛋白在感染早期的显著协调和富集。晚期以细胞蛋白修饰、膜蛋白复合物形成和细胞死亡诱导为特征。这些结果,连同表型观察,提供了进一步的洞察分子机制的马铃薯害虫抗性。

材料:马铃薯(cv. Sarpo Mira)、疟原虫分离物包被的游动孢子悬浮液、TMT试剂(Thermo Fisher Scientific,San Jose,CA,USA)
目的:蛋白质组学已经成为分子分析的一种可行的替代方法,为更好地理解植物与病原的关系提供了信息和工具。近年来,蛋白质组学在寻找候选蛋白的大规模功能分配方面发生了巨大的发展,利用这种方法,已经鉴定出几种在马铃薯-马铃薯互作过程中表达的蛋白质。基于双向电泳(2-DE)的蛋白质组学、基于凝胶的蛋白质SHOTGUN大规模质谱鉴定分析和最近的非标记定量蛋白质组学技术为我们理解马铃薯和马铃薯之间的相容和不相容相互作用提供了线索。本研究采用TMT法对马铃薯的蛋白质丰度变化进行了测定和比较。在三个关键时间点叶面施用虫草游动孢子悬浮液,覆盖马铃薯-虫草卵菌丝体相互作用的早期(EI)和晚期(LI)。共鉴定出1229个差异表达蛋白(DEP),其中早期75个,晚期723个。早期和晚期鉴定的蛋白在病原菌的早期识别、信号转导、抗病过程以及可能的发病机制中起着重要作用。本研究将有助于进一步了解马铃薯与害虫相互作用的分子机制。

技术路线分析:
实验:马铃薯品种Sarpo Mira在温室中生长,控制条件为20℃、16:8光-暗循环和70%相对湿度。将5周龄的植株转移到100%湿度和10:14光暗循环的感染室中。6h后,用侵染疟原虫分离物包被的游动孢子悬浮液喷洒植株,直到叶表面完全浸透游动孢子悬浮液(15000个孢子囊/mL)。根据规定,在接种后0、48和120h收集样本,并分别标记为对照、EI和LI。接种后立即采集0 dpi样本,接触时间小于1分钟。接种后两天,相对湿度保持在100%,其余时间调整到90%。在每个时间点,从三个独立的生物学实验中采集完全展开的上部叶片样本。所有材料均在液氮中冷冻,并在-80°C下储存,直至使用。接着进行蛋白质提取工作,得到一定浓度的蛋白质裂解液;然后将该样品的裂解液进行酶解,并在酶解后中的滤液中滤液中收集肽,用波长为280nm(OD280)的光密度测定肽的浓度;最后根据制造商的推荐,使用TMT试剂标记每个样品,使用LC-MS/MS进行相关质谱分析。

结果与讨论:
1. 疾病三个阶段的分段表型差异
图1a,b所示为用侵染疟原虫激发的Sarpo Mira叶片表型的时间序列评估。在这些试验中,每种处理使用三个复制品。在接种hpi后0小时(对照组)向整个马铃薯植株喷洒侵染芽孢杆菌(P.infestans zoospores)后,未观察到明显的超敏反应(HR)损伤;然而,在48hpi(EI)时,出现HR损伤,导致局部坏死,类似于侵染芽孢杆菌引起的超敏反应,在120hpi时(LI)叶片发生了更大的H-诱导的坏死,与典型的R基因介导的HR损伤扩展一致,如之前在Sarpo Mira中报道的。如图1b所示,LI相对于EI的病变大小存在显著差异,与之前的研究一致。根据这些结果,我们将控制时间点和EI之间的间隔指定为早期疾病阶段,将EI到LI之间的时间点指定为晚期疾病阶段。

图1:三个时间点的表型观察。对照组对应接种后0小时的时间点,EI对应48hpi,LI对应120hpi。(a) 不同暴露时间虫害游动孢子引起的损伤大小。接种后第2天和第5天分别测量各病灶直径(b)。在这些试验中,每种治疗方法使用三个复制品。条形图表示三个副本的标准偏差。采用t检验分析统计学意义。星号表示差异显著(p<0.05)。
2. 马铃薯卵菌侵染叶片蛋白质表达的研究进展
为了进一步了解晚疫病早期和晚期蛋白质丰度的变化,我们在3个关键时间点(对照时间点、EI时间点和LI时间点)用TMT标记进行了比较蛋白质组调查。同时收集了3个生物复制品。通过这些测量,在时间序列分析中共鉴定了15813个优质肽(每个时间点至少两个重复),对应于4643个蛋白质。在4643个蛋白质中,1229个(至少一个独特的肽)在两个时间点的成对比较中被发现是差异表达蛋白质(DEP)。在1229个DEP中,952个具有功能注释。
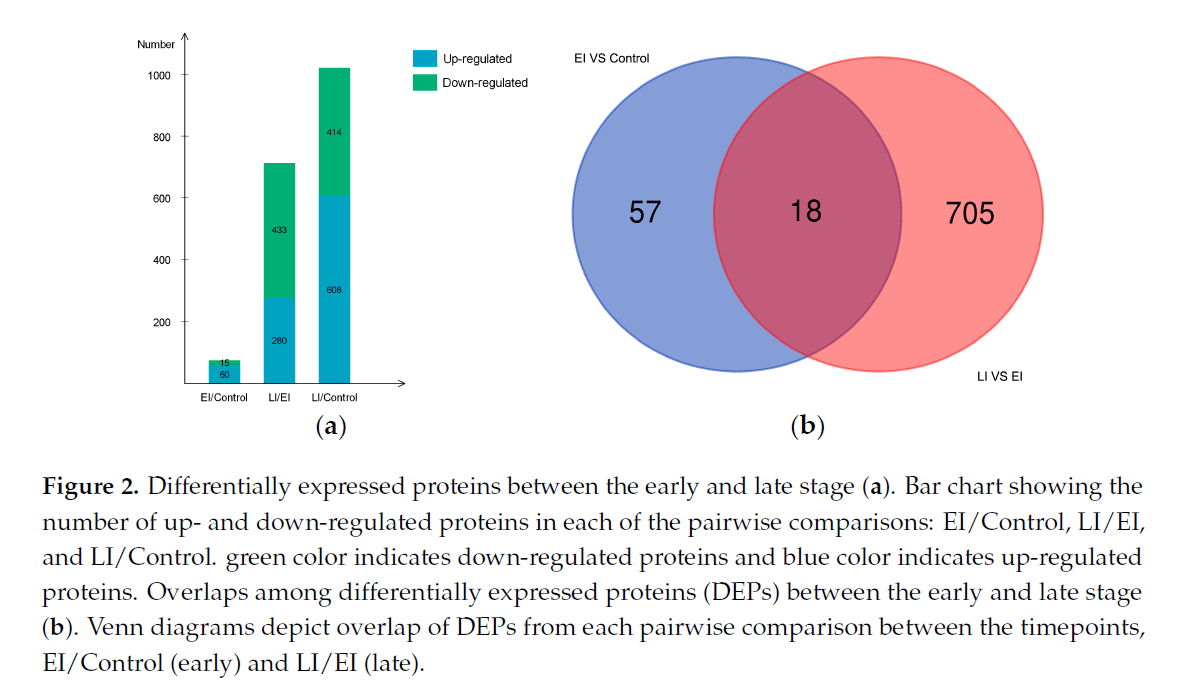
图2:早期和晚期差异表达蛋白(a)。条形图显示每对比较中上调和下调蛋白质的数量:EI/对照、LI/EI和LI/对照。绿色表示下调蛋白,蓝色表示上调蛋白。差异表达蛋白(DEPs)在早期和晚期(b)之间的重叠。Venn图描述了时间点EI/控制(早期)和LI/EI(晚期)之间每对比较的DEP重叠。
3. 差异表达蛋白的基因本体论(GO)分类
利用基因本体论分类和KEGG分析揭示了本研究中发现的所有DEPs的含义(图3a)。基因本体分析表明,DEPs中有大量的生物过程范畴。蛋白质类别“代谢过程”(41.3%)、细胞过程(32%)和“刺激反应”(6.1%)是最丰富的类别(图3a)。
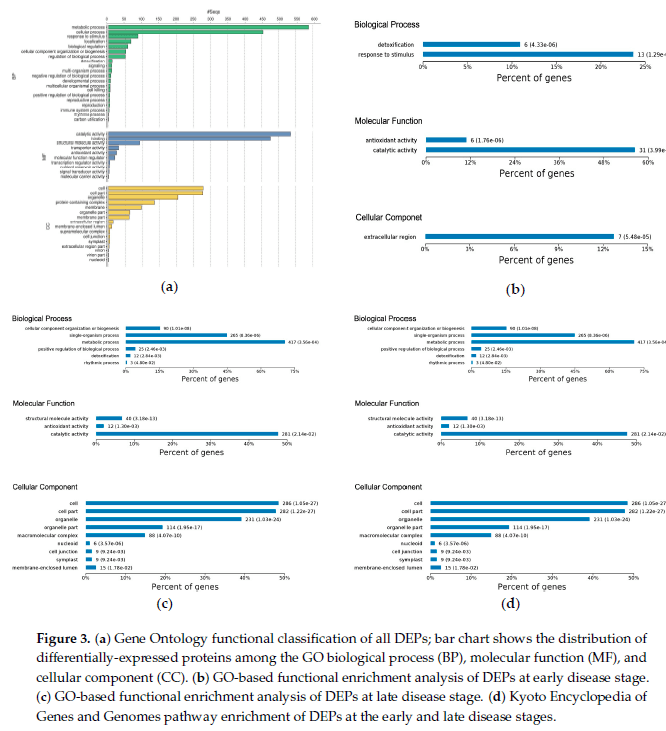
图3:(a) 所有DEPs的基因本体功能分类;条形图显示GO生物过程(BP)、分子功能(MF)和细胞成分(CC)中差异表达蛋白的分布。(b) 基于GO的DEPs早期功能富集分析。(c) 基于GO的疾病晚期DEPs功能富集分析。(d) 京都百科全书基因和基因组途径在疾病早期和晚期的DEPs富集。
利用KEGG数据库进行的途径覆盖分析发现,苯丙酸生物合成途径在疾病早期最显著地富集(图3d)。这些结果表明,所鉴定的蛋白质代表了整个蛋白质组的功能活性亚组,该亚组与马铃薯对感染卵子杆菌的反应有关。
4. 参与早期和晚期疾病的蛋白质的差异表达模式
早期疾病反应蛋白:75种蛋白质在疾病过程的早期阶段显示出蛋白质丰度的显著差异(EI/对照比较蛋白质),其中60种DEP上调,15种DEP下调(图2a);其表达谱如图4所示。GO富集试验结果(假发现率<0.05,p<0.01)显示,这些蛋白质主要与解毒和对刺激的反应有关,包括应激反应、防御反应、氧化爆发和细胞分解代谢过程(图3b)。

图4:疾病早期差异表达蛋白的层次聚类分析。显示蛋白质表达变化的热图:高表达水平的蛋白质(红色);低表达水平的蛋白质(蓝色)。
晚期疾病反应蛋白:图5显示了后期确定的723个DEP的剖面图。在该组中,共有280个DEP上调,443个DEP下调,如图2a所示。功能富集表明,大多数蛋白质参与细胞成分组织、代谢过程和单有机体过程,包括细胞蛋白修饰过程和膜蛋白复合物形成(图3c)。
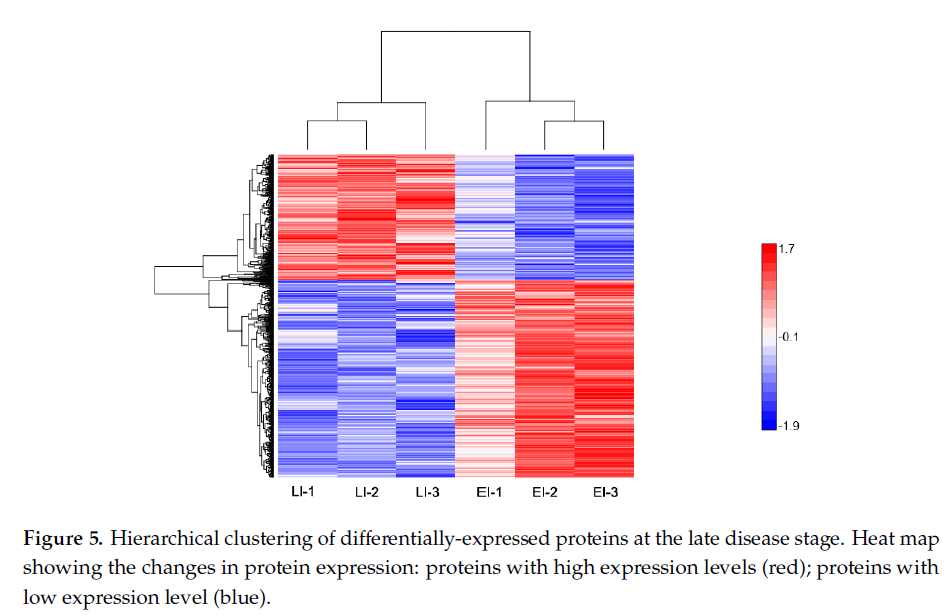
图5:疾病晚期差异表达蛋白的层次聚类分析。显示蛋白质表达变化的热图:高表达水平的蛋白质(红色);低表达水平的蛋白质(蓝色)。
常见共享蛋白在疾病早期和晚期的差异调节:我们的比较分析确定了在EI/对照比较和LI/EI比较之间共享的18种蛋白质(图2b)。我们推断,这些共同的蛋白质组可能对控制从基于PAMPs(PTI)的一般植物防御诱导转变为效应器触发免疫(ETI)的相同信号作出反应,并且可能具有类似的表达模式。事实上,在18种常见的dep中,有10种在整个时间过程中表现出相似的表达谱。根据GO分析,这10种蛋白主要位于细胞外区域、细胞膜和含蛋白复合物内,参与能量生产、囊泡介导的转运和氧化应激反应。M1CUM0具有与萜类生物合成相关的结构域功能,部分抗真菌植物抗毒素在病原菌入侵时限制菌丝生长。
5. 早期和晚期疾病阶段差异蛋白的验证
为了验证时间序列蛋白质组学实验中的DEPs,共选择了7种蛋白质,其中4种是从早期疾病阶段随机选择的,其余3种是从晚期选择的,通过western blot分析来验证表达水平(图6)。western blot结果与时间序列蛋白质组学数据一致,有力地支持了本文结果的可靠性。
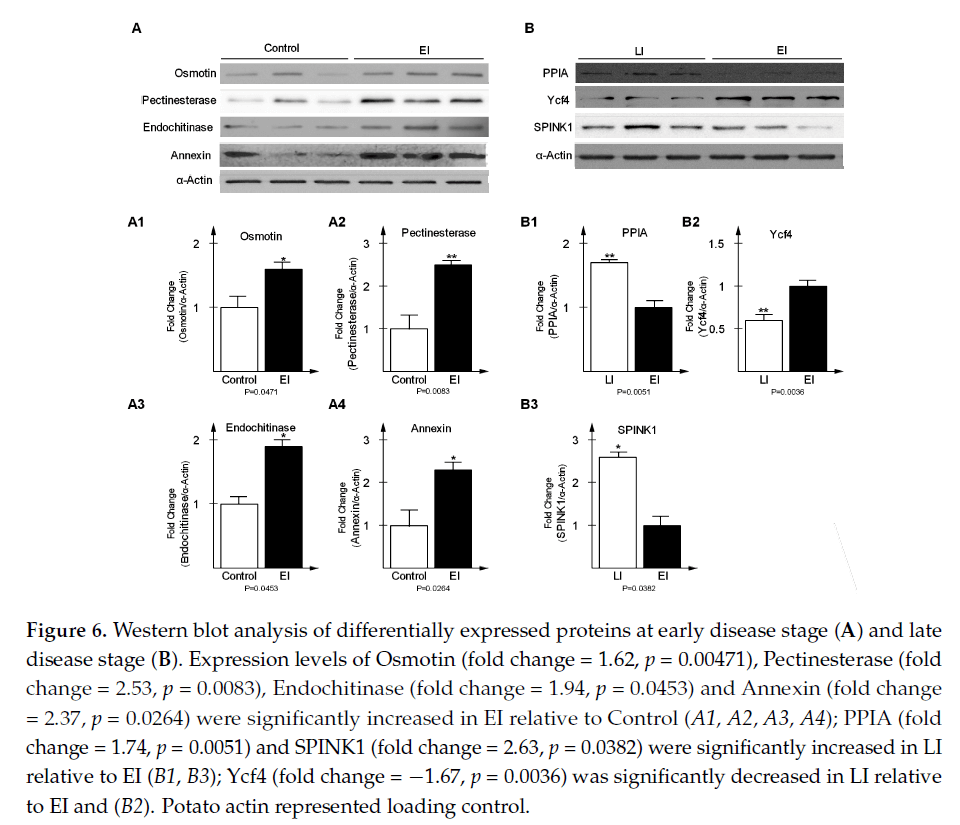
图6:早期(A)和晚期(B)差异表达蛋白的Western blot分析。与对照组(A1、A2、A3、A4)相比,EI组Osmotin(fold change=1.62,p=0.00471)、果胶酯酶(fold change=2.53,p=0.0083)、内切酶(fold change=1.94,p=0.0453)和Annexin(fold change=2.37,p=0.0264)表达水平显著升高(fold change=1.74,p=0.0051),SPINK1(fold change=2.63,p与EI(B1,B3)相比,LI显著增加了=0.0382;与EI和(B2)相比,LI显著减少了Ycf4(倍数变化=-1.67,p=0.0036)。马铃薯肌动蛋白代表负荷控制。
6. 疾病早期和晚期的必需蛋白质
cv.Sarpo Mira感染早期阶段的蛋白-蛋白相互作用。为了揭示感染疟原虫入侵早期蛋白质之间的功能相互作用,我们通过检索细胞景观中相互作用基因/蛋白质的搜索工具(https://string-db.org)分析了EI/对照比较中的75个DEP(图7a)。

图7:在(a)感染早期和(b)感染晚期蛋白质显著变化的网络分析结果。
在本研究中,这几种渗透因子(M0ZTM9,FC=1.34)从对照组到EI,再到LI的显著上调,表明细胞壁内的致病相关蛋白之间存在显著的协同作用,从而促进基础抵抗反应。总之,这些结果表明,在疾病感染的早期阶段,防御库有一个协调一致的上调调节,如病原体识别蛋白、信号分子、抗菌化合物、细胞运输,这些都是启动对侵染疟原虫广泛抗性的基础。cv.Sarpo Mira感染后期的蛋白-蛋白相互作用我们分析了在LI/EI比较中发现的DEPs的蛋白质相互作用,并发现与网络中几个hub蛋白质的多因素相互作用(图7b)。感染疟原虫病过程的晚期通常以定植和坏死为特征;在这一阶段,细胞死亡调节蛋白(M1D3S7、M1CJS7、M1BAS6)之间观察到了强的结构域自相互作用。
7.蛋白质表达与mRNA的实时qPCR相关性分析
为了评价mRNA与蛋白质水平的相关性,我们用RT-qPCR分析了编码8种代表性蛋白质的基因在3个时间点的相对表达模式。在疾病感染的早期阶段,8个基因中有3个编码参与细胞壁结构修饰的蛋白质,其表达水平较高(图8a)。在感染周期的后期,对其余五个基因观察到了类似的模式(图8b)。这些结果证实了蛋白质组分析中所观察到的编码蛋白水平的增加,并表明这些蛋白的丰度可能在转录水平上受到调节。

图8:实时多聚酶链反应(PCR)定量分析8种差异表达蛋白和mRNA在疾病早期(a)和晚期(b)的表达。绿色条表示由串联质谱标记(TMT)确定的表达水平,橙色条表示mRNA的相关表达。所有数据均表示为平均值±标准差(每个时间点n=3)。
结论:本研究揭示了马铃薯卵菌感染早期和晚期蛋白质丰度的变化及蛋白质的生理作用。我们还推断了在两个疾病阶段发生的蛋白质相互作用,或者是通过实验验证的物理相互作用,或者是进一步了解疾病过程的预测。总的来说,我们的分析表明,在感染早期鉴定的差异表达蛋白在信号转导和基础防御反应中发挥了重要作用,而晚期疾病过程的特征是与抗病过程和细胞死亡相关的R蛋白显著丰富。然而,疾病晚期的一些蛋白质可能与疾病晚期的易感过程有关。因此,本文报道的数据对于进一步研究马铃薯晚疫病抗性过程中可能涉及的机制具有重要的实用价值。