
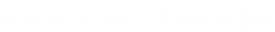
文献解读
项目论文|云南野生稻对褐飞虱特异抗性的iTRAQ比较蛋白质组学研究-BMC Plant Biology-201901
在各种害虫中,危害水稻的褐飞虱(BPH)是危害水稻的主要害虫。了解抗药性机制是有效控制BPH的关键一步。本研究旨在探讨水稻对褐飞虱(BPH)的抗性(PR)、感病(PS)和杂交种(HR)与褐飞虱(BPH)的互作蛋白质组学,以了解水稻对褐飞虱的抗性机制。

结果:通过iTRAQ蛋白质组学研究,从这三个水稻品种中鉴定出4900多个蛋白质。BPH感染后,PR、PS和HR分别检测到414、425和470个差异表达蛋白(DEPs)。已鉴定的DEPs主要富集在与次生代谢产物的生物合成、碳代谢、乙醛酸盐和二元酸盐代谢有关的类别中。一种双组分反应调节蛋白(ORR22)可能参与BPH感染后的早期信号转导。在抗性水稻品种(PR)中,6个DEPs,即两个脂氧合酶(LOXs)、一个脂肪酶、两个直接蛋白(dirigent proteins,DIRs)和一个Ent-cassa-12,15-二烯合酶(OsDTC1)与遗传性BPH抗性有关。热休克蛋白(HSP20)可能参与水稻对BPH侵染的生理反应,成为水稻分子标记辅助选择(MAS)的潜在靶点。定量实时聚合酶链反应(qRT-PCR)显示8个编码BPH抗性相关代谢蛋白的基因。在籽粒发育过程中,这些基因的表达在转录和翻译水平上发生了变化。这项研究提供了全面的细节关键蛋白质在兼容和不兼容的交互在BPH侵扰,这将有助于进一步研究水稻抗褐飞虱的分子基础和育种BPH-resistant水稻品种。

材料:BPH(褐飞虱)、水稻、iTRAQ蛋白质组学
目的:利用iTRAQ蛋白质组学分析BPH侵染水稻,进一步了解褐飞虱侵染过程中复杂的分子和细胞事件,褐飞虱抗性水稻等。本研究共分为6个处理,即有(PR)BPH感染和无(PR)BPH感染的水稻;有(PS)BPH感染和无(PS)BPH感染的水稻;以及有(HR)BPH感染和无(HR)BPH感染的水稻。植物一直生长到孕穗期,当BPH被引入三个感染处理。在BPH感染开始后30天收集茎组织。选择视觉健康的植物进行无BPH感染的治疗。对于BPH感染的处理,选择有严重感染迹象的植物进行采样。每次治疗收集三个独立的复制品。
结果:
水稻褐飞虱侵染的表型研究:继侵染后,BPH引起幼苗萎蔫,首先导致感病水稻品系PS出现稻飞虱灼伤症状,其次是HR和PR(两个抗BPH品系)(图1b)。叶片的表观损伤以PR最低,HR居中,PS最高。感病(PS_B)和抗性(PR_B和HR_B)水稻品系的表型差异非常明显,与不同抗性水稻品种感染BPH后的生理表型结果一致。

图1:褐飞虱接种不同水稻基因型的发生与症状。稻田褐飞虱为害及典型症状见右下角。褐飞虱接种水稻基因型的表型研究。杂交后代褐飞虱分别接种(HRúB)和不接种(HR)褐飞虱;杂交后代褐飞虱、药用植物褐飞虱和水稻褐飞虱分别接种(HRúB、PRúB、PSúB)和不接种害虫的褐飞虱(HR、PR、PS)
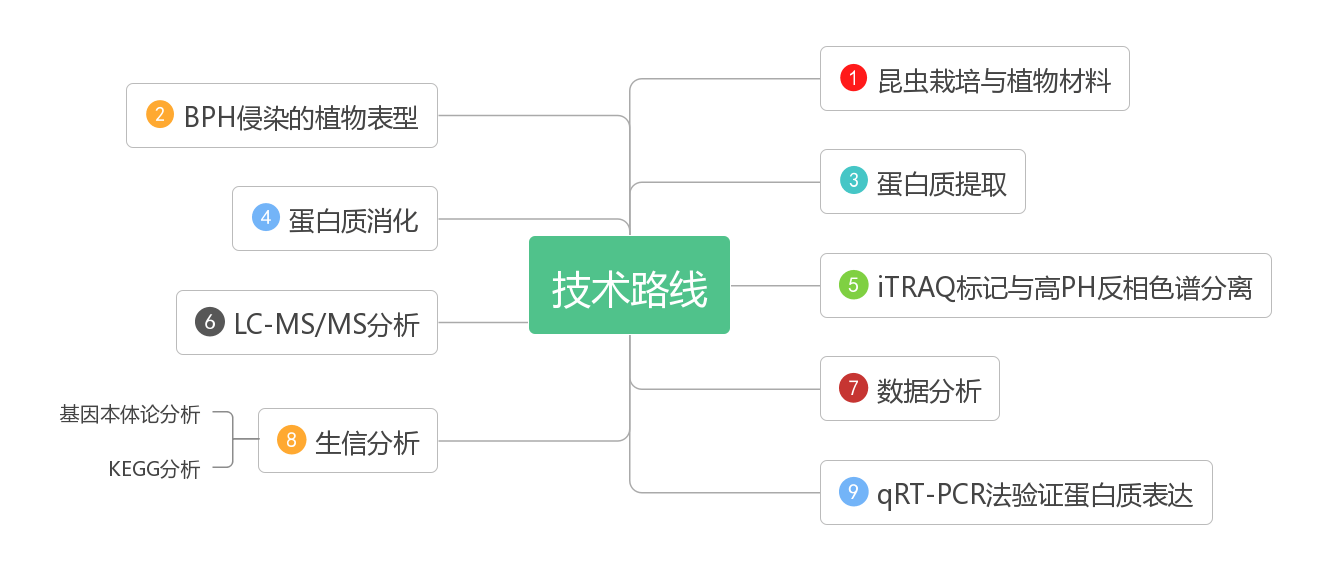
三个品种接种BPH后差异表达蛋白的研究:通过iTRAQ分析,从这三个品种中共鉴定出4907个蛋白质。在PR和PS之间,共鉴定出462个差异表达蛋白(DEPs),其中抗性PR增加232个,抗性PR减少230个,与PS相比,HR增加281个,下降237个,说明这些水稻品种幼苗蛋白质表达差异较大。接种BPH后,三个水稻品种的蛋白质表达均发生显著变化:PR中检测到414个DEPs,其中200个表达水平上调,其余214个表达水平下调;PS中检测到423个DEPs,接种后248例上调;接种后470例DEP中190例上调(图2)。
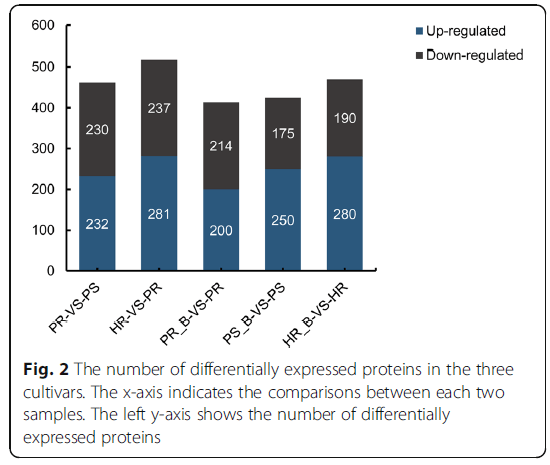 图2:三个品种差异表达蛋白的数量。x轴表示两个样本之间的比较。左y轴显示差异表达蛋白的数量
图2:三个品种差异表达蛋白的数量。x轴表示两个样本之间的比较。左y轴显示差异表达蛋白的数量
接种BPH后,三个水稻品种对BPH侵染的反应产生1084个DEPs(328 + 59 + 21 + 62 + 60 + 280 + 274),如韦恩图(图3a)所示。HRúB与HR、PSúB与PS、PRúB与PR共享的21个DEP可赋予对BPH的潜在广谱抗性。存在这样一种可能性,即HRúB vs.HR和PRúB vs.PR共享的59个DEPs与水稻对BPH的主动抗性有关。由于本研究的一个主要目的是筛选与BPH抗性相关的内部遗传蛋白生物标记物,我们选择了在背景和BPH感染植物之间一致差异表达的蛋白质。我们发现,4个比较组HR-B vs.HR、PR-B vs.PR、PR-PS和HR-PS共有15个DEP(图3b),其中只有一个(热休克蛋白HSP20,B0FFN6)在BPH感染后的每次比较中显著上调。这些结果表明抗性品种与敏感品种之间存在差异,进一步分析这些基因可能有助于揭示抗性机理。
图3:抗感水稻品种接种BPH后DEPs的韦恩图。HR-B-VS-HR、PS-B-VS-PS和PR-B-VS-PR之间比较的韦恩图。HR-B-VS-HR、PS-B-VS-PS、PR-B-VS-PR、PR-VS-PS和HR-VS-PS之间比较的韦恩图
为了获得BHP感染后蛋白质组变化的综合表征,所有1084个DEP均使用基因本体论术语进行注释并进行基因本体论功能分析。以这些DEPs为代表的生物过程主要有代谢过程、刺激反应、细胞过程和单生物体过程。根据其分子功能性质,这些蛋白质主要分为催化活性、结合活性、结构分子活性、电子载体活性、转运活性、抗氧化活性和营养贮藏活性(图4)。利用KEGG数据库对这些DEPs进行了进一步的研究,发现它们在次生代谢产物(10.7%)、核糖体(5.0%)、碳代谢(3.6%)、糖基和二羧酸代谢(3.2%)、卟啉和叶绿素代谢(2.8%)、光合作用(1.8%)、过氧化物酶体(1.8%)的生物合成中有丰富的表达,光合作用生物的碳固定(1.7%)、氮代谢(1.1%)和花青素生物合成(0.4%)(图5)。
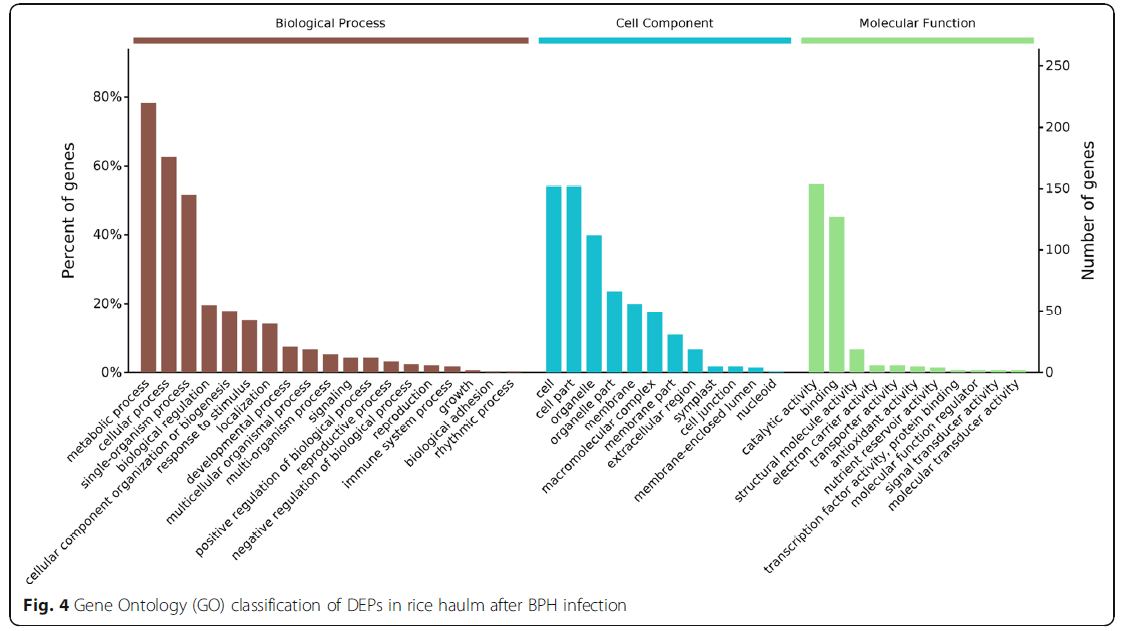
图4:BPH感染后稻谷中DEPs的基因本体论分类
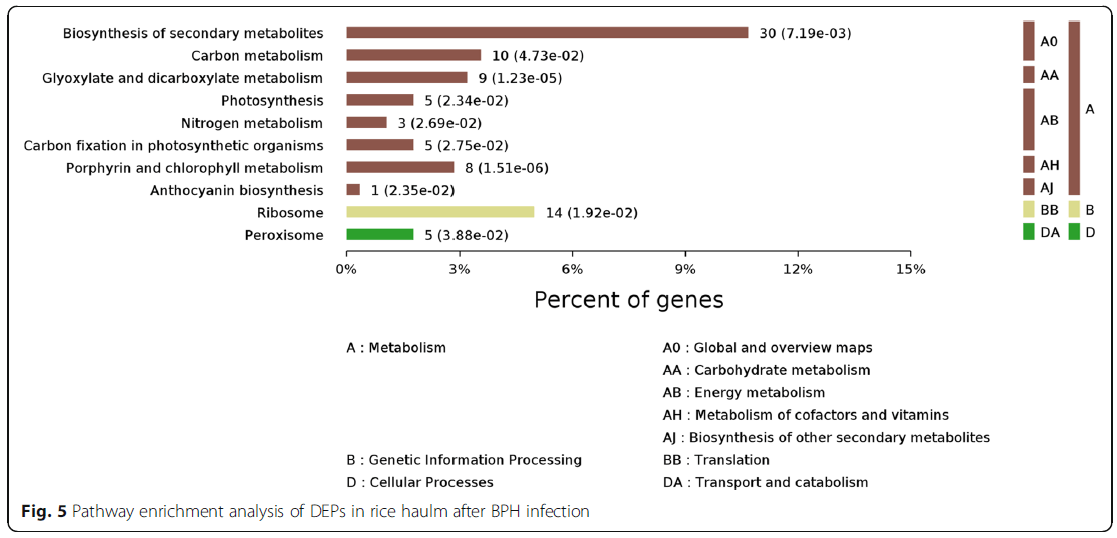
图5:BPH感染后稻谷中DEPs的途径富集分析
定量RT-PCR验证:为了验证mRNA及其蛋白表达模式之间相关性的定量结果,我们随机选择了8种蛋白,采用定量RT-PCR(Q-PCR)分析方法对其进行动态转录表达谱的评估。图6显示编码Q7FAS1基因的mRNA表达模式与蛋白质表达模式相似。PS_B-VS-PS、HR_B-VS-HR和PR_B-VS-PR组的Q6AVH9、Q6K832和Q6Z7B3基因在mRNA和相应蛋白之间的表达谱相似。另一方面,Q2R1U4、Q6YUV3、Q7XRT6和Q8H7X8基因的mRNA表达模式与相关蛋白的表达模式相反,这可能是翻译或翻译后修饰的结果。
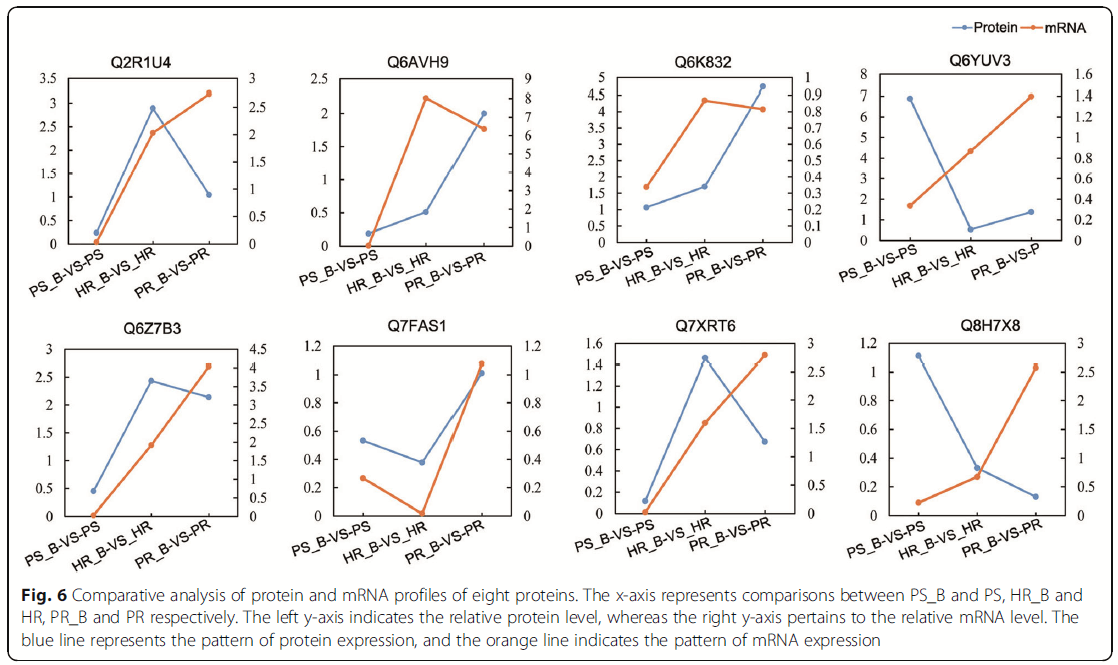
图6:8种蛋白质的蛋白质和mRNA谱的比较分析。x轴分别表示PS_B和PS、HR_B和HR、PR_B和PR之间的比较。左y轴表示相对蛋白水平,而右y轴表示相对mRNA水平。蓝线代表蛋白质的表达模式,橙色线代表mRNA的表达模式
讨论:褐飞虱是水稻的主要害虫之一,对水稻的大规模生产构成严重威胁。培育抗病品种是提高作物生产性能和控制农业害虫最有效、最环保的途径。此外,野生稻属植物具有丰富的遗传多样性,实际上尚未开发利用,因此可以作为抗BPH的关键来源。目前已有19个以上的抗BPH基因被鉴定为具有BPH抗性,并通过QTL定位被分配到栽培稻和野生稻的染色体上。QTL在标记辅助植物育种中经常被用来预测表型。本研究采用iTRAQ比较蛋白质组学方法,对野生栽培抗BPH水稻品系PR和抗BPH水稻品系PS中的差异积累蛋白进行了鉴定,其抗BPH杂交品系HR有助于了解BPH与水稻之间的分子互作关系,以及水稻对BPH的遗传抗性。与以往基于传统二维蛋白质组学的研究相比,DEPs的数量显著增加。
BPH感染后参与早期信号转导的蛋白质:
激素信号通路在水稻防御信号网络中起着重要作用。水稻对BPH的防御和植物激素的作用是相当复杂的,并且在不同基因型间存在差异。BPH的入侵通常会提高水稻中乙烯(Et)、水杨酸(SA)和茉莉酸(jasmonic acid)的产量。本研究鉴定了BPH接种后HR-B组与HR组、PS-B组与PS组、PR-B组与PR组共21个DEPs。这些DEPs可能参与水稻与BPH的早期相互作用。且两组分反应调节因子(ORR22)在BPH感染后显著上调,在H、M和W分别为2.61、9.86和1.64,提示ORR22可能在BPH的抗性中起关键作用。
水稻抗褐飞虱遗传抗性的相关蛋白:
通常,病原相关蛋白与植物防御过程直接相关,并受到病原/寄生虫攻击的刺激。在相容和不相容的相互作用中,抗性的可持续性取决于这些蛋白质的相互作用以及它们积累或激活植物防御系统的方式。为了帮助了解水稻对褐飞虱的遗传抗性,我们鉴定了两个抗性品种(HR和PR)在褐飞虱侵染后积累的59个DEP。利用Swissprot数据库对这59个DEPs进行了注释分析,结果表明,6个与水稻与BPH互作有关的DEPs在两个抗性品种中均上调。这六种dep被认为与BPH的遗传抗性有关(图7)。
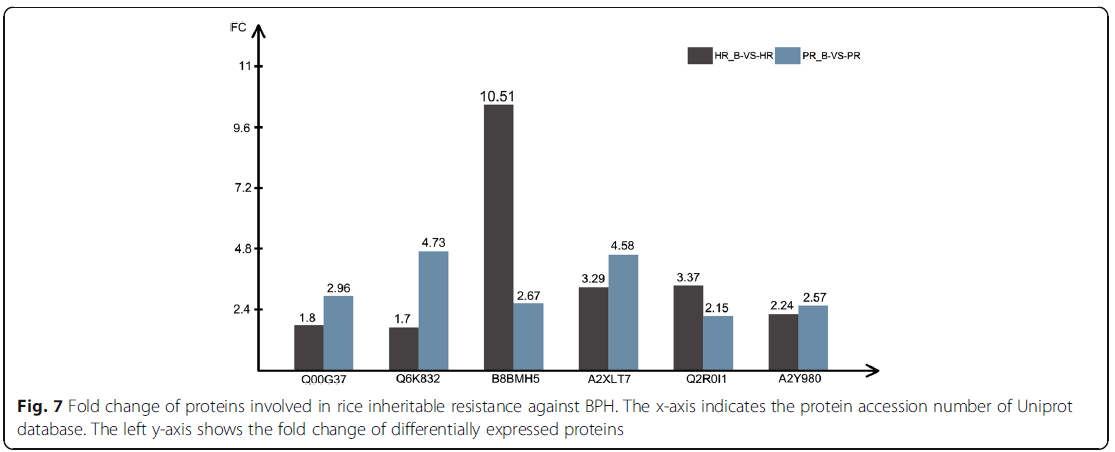
图7:水稻抗褐飞虱遗传抗性相关蛋白的折叠变化。x轴表示Uniprot数据库的蛋白质登录号。左y轴显示差异表达蛋白的折叠变化
一种复杂的防御机制已经在植物中发展起来,它可以利用多种防御途径的有组织的作用来对抗各种非生物和生物胁迫。植物防御反应涉及多种有机化合物,如氧化脂质、乙烯、水杨酸等。氧化脂质是由多不饱和脂肪酸氧化修饰而产生的一类化合物,起着植物信使的作用。除各种发育过程外,氧化脂质还参与介导作物对非生物和生物胁迫的防御反应。氧化脂质的生物合成是通过多不饱和脂肪酸氧化合成脂肪酸氢过氧化物而开始的。为了确定第一步,脂肪酶在不同的氧合酶生物合成途径中与单独的脂氧合酶(lox)协同工作。近年来对水稻LOX的研究表明,LOX的上调可能是介导JA爆发、JA与SA间串扰和抗性权衡的主要节点。在本研究中,发现BPH感染后HR和PR中有两种LOX(A2XLT7和B8BMH5)和一种脂肪酶(Q6K832)上调,这与LOX参与了害虫诱导的JA生物合成的观点相一致,在控制水稻对以咀嚼和韧皮部为食的草食动物的抗性中起着关键作用。本研究发现,BPH感染后HR和PR中有两种DIRs(A2Y980和Q2R0I1)表达上调。这可能表明,在BPH感染时,DIR组分的增加促进了木脂素及其相关次级代谢产物的含量,这些物质可能是干扰BPH生长的物质。Ent-cassa-12,15-二烯合酶(OsDTC1)在植物cassanes生物合成中具有重要作用。BPH感染后HR和PS中OsDTC1均上调,这可能有助于提高植物cassanes的水平,促进BPH的抗性。
抗BPH水稻育种的潜在标记蛋白:
开发抗病品种被认为是保护水稻免受褐飞虱侵害的一种有效和对环境负责的方法。传统育种方法受到遗传复杂性、产量构成因素遗传方差低、选择方法效率低、环境变异性大、基因型与环境交互作用强等因素的限制。基本上,基于组学的一些方法提高了我们确定目标基因成分和代谢途径的能力,这些基因成分和代谢途径控制着特定的性状,因此使我们能够通过筛选和分析平台支持选择策略。在这项研究中,我们发现15个DEP由HR与HR,PR与PR,PR与PS和HR与PS的所有配对共享,其中只有一个(热休克蛋白HSP20,B0FFN6)在BPH感染后显著上调。由于各种胁迫条件,小分子热休克蛋白(sHSPs)/HSP20被诱导并在植物抵御生物和非生物胁迫中发挥重要作用。最近的一项研究表明,水稻的小HSP20受到RSV感染的显著影响,RSV感染是通过小褐飞虱(SBPH)的活性以持续循环的方式传播的。我们假设HSP20可能参与BPH侵染的生理优势反应,使其成为标记辅助选择(MAS)的潜在靶点,从而显著提高育种抗BPH水稻的效率。
结论:综合蛋白质组学分析和qRT-PCR分析表明,褐飞虱入侵导致抗褐飞虱和感褐飞虱水稻品种的蛋白质发生复杂变化。本研究结果为进一步了解褐飞虱侵染过程中复杂的分子和细胞事件提供了新的线索,为褐飞虱抗性水稻的育种提供了潜在的有用工具。ORR22的激活是抗BPH和感病水稻对BPH侵染的共同反应,它可能通过持续促进SA在早期信号转导中发挥抗BPH的作用。重要的是,抗病品种和感病品种对褐飞虱的遗传抗性存在显著差异,抗性水稻对褐飞虱的侵染表现出强烈的反应,表现为蛋白质的数量和变化程度。LOXs、DIRs和OsDTC1是BPH遗传抗性的关键酶。此外,我们发现HSP20(热休克蛋白)可能是BPH抗性育种的潜在靶点。