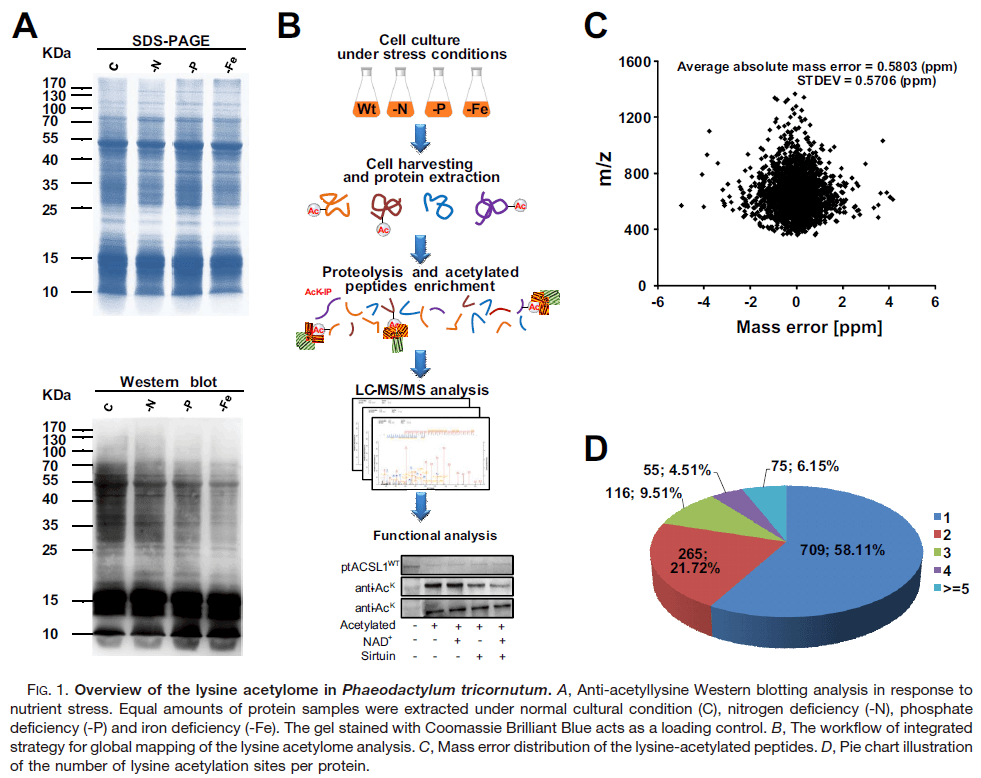
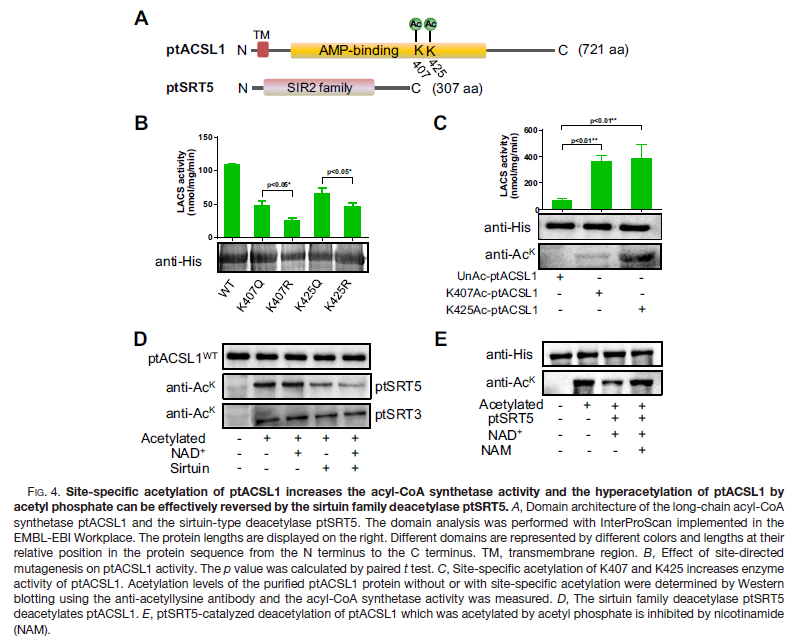
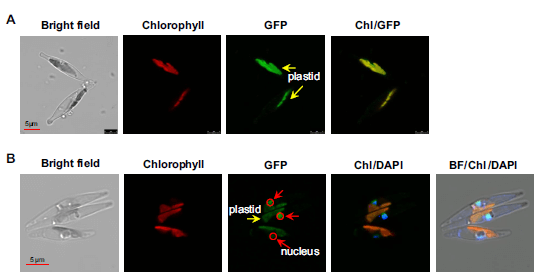
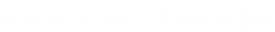LACS定点突变的活性实验。将ptACSL1的位于AMP结合区的K407和K425分别突变为谷氨酰胺(K407Q,K425Q),谷氨酰胺(K407Q,K425Q)。酶活结果表明所有的定点突变都导致LACS活性下降(图4B)。通过大肠杆菌重组纯化K407-,K425-乙酰化,和未乙酰化的ptACSL1,酶活发现与未乙酰化的ptACSL1相比,K407和K425的乙酰化导致ptACSL1的LACS活性显著增加(图4C)。
Sirtuin是ptACSL1的一种常见的去乙酰化酶。纯化Sirtuin(ptSRT5)并于乙酰化的ptACSL1进行孵育,发现ptACSL1在NAD+存在下能有效地脱乙酰,而ptSRT3在同一反应体系中不能诱导ptACSL1脱乙酰(图4D)。如果添加烟酰胺(SIRT脱乙酰化酶家族的特异性抑制剂)可完全抑制ptSRT5对乙酰化ptACSL1的脱乙酰化活性。以上表明AcP乙酰化的ptACSL1可以被三角褐指藻中SIRT脱乙酰酶家族的sirtuin蛋白ptSRT5逆转。